
Episode 217: Adverse Childhood Experiences - HPA axis & Brain changes: cortisol, amygdala, hippocampus, cytokines, & epigenetics (Part 3 of ACE series)
By listening to this episode, you can earn 1.5 Psychiatry CME Credits.
Other Places to listen: iTunes, Spotify
Article Authors: Liam Browning, David Puder MD
Peer reviewed by Erica Vega, Joanie Burns
None of the authors or presenters have any conflicts of interest to report.
In this week’s episode, we continue our series on Adverse Childhood Experiences by delving deeper into the lasting effect of ACEs on the brain and body. We explore the intricate impact of ACEs on the HPA axis, inflammation, and neurobiology, shedding light on their role in various psychiatric disorders. We highlight how these changes may indicate a shared phenotype resulting from early adversity but that they likely do not explain the entire effect ACEs have on an individual.
Highlights
ACEs lead to a more blunted cortisol response to stress
ACEs may lead to epigenetic changes in glucocorticoid receptor genes
ACEs are associated with morphological changes in the brain, including decreased hippocampal volume and decreased white matter integrity in the corpus callosum
ACEs increase levels of chronic inflammatory cytokines
ACEs increase amygdala reactivity to negative emotional stimuli and reduce the activity of the reward system during reward anticipation
What Are The Biological Mechanisms Through Which ACEs Could Exert Their Effects?
The mass of research in psychiatry over recent years has been directed towards understanding the biological underpinnings of disorders in hopes of providing diagnostic clarity and targeted treatment.
Researchers have begun to recognize the implications of Adverse Childhood Experiences (ACEs) on mental and physical health outcomes (see episode 1 and 2 of our series), which has led to thousands of studies aimed at understanding the biological effects of ACEs.
Many of these studies are based on the hypothesis that recurrent stressful experiences during development can disrupt the body's stress response system, particularly the glucocorticoid system, also known as the hypothalamic-pituitary-adrenal (HPA) axis. If the HPA-axis is highly active while it is undergoing development due to chronic stressful experiences, there may be a long-term upregulation or downregulation of its functionality. It’s hypothesized that these change in long-term functioning could be mediating by epigenetic modifications of genes involved in HPA-axis function.
Some researchers also believe that, given the glucocorticoid system’s broad functionality, particularly in neurodevelopment, elevated glucocorticoid levels during sensitive periods of brain development can cause widespread deficits in neurodevelopment, leading to increased risk for psychiatric disorders.
Cortisol And The Glucocorticoid System: Childhood Trauma Causes A More Blunted Cortisol Response To Stressors
Section Highlights:
Childhood maltreatment causes a blunted cortisol response to stress in lab-based settings and to dexamethasone in adults.
A blunted cortisol response is associated with other psychiatric disorders, including PTSD, BPD, and atypical depression, and is correlated with adiposity, inflammation, and poor self-rated health.
Childhood maltreatment may lead to hypocortisolism by decreased morning wakeup cortisol levels in children, leading to flatter daily cortisol rhythms. However, some studies show basal cortisol is increased in abused or neglected children.
What is the function of the HPA-axis?
The canonical pathway of the glucocorticoid system begins with the detection of a threat and the activation of corticolimbic regions, primarily the PFC, amygdala, and hippocampus. These regions indirectly (via the stria terminalis) activate neurons in the paraventricular nucleus (PVN) of the hypothalamus carrying corticotropin releasing hormone (CRH), causing the release of CRH into the hypophyseal portal system. CRH travels to the anterior pituitary where it acts on CRHR1 (CRF1 in animals) to cause the release of ACTH into the bloodstream. ACTH then travels to the adrenals, which then produce cortisol (corticosterone in rodents). Cortisol can bind to glucocorticoid receptors within the cell. Once bound with cortisol, the glucocorticoid receptor can translocate to the nucleus where it changes transcription of a variety of genes involved in various functions. Cortisol helps mobilize glucose for energy, ramp up the immune system, enhance salt retention, and modulate the autonomic nervous system.
Once the acute stressor has passed, cortisol acts on glucocorticoid and mineralocorticoid receptors (GR and MR) in the hippocampus, PFC, hypothalamus, and anterior pituitary to suppress the HPA-axis (Urlich-Lai and Patterson, 2009).
Cortisol also can act on G-protein coupled receptors on the surface of neurons and glial cells to influence second messenger systems associated with neurotransmitters. These effects are thought to mediate glucocorticoids' ability to enhance the consolidation of memories associated with the acute stressor (Roozendaal, 2009).
Aside from increasing during the acute stress response, cortisol increases in pulsatile rhythms in a diurnal pattern. Cortisol levels increase 30-45 minutes prior to waking and continue to increase until a peak about 30 minutes after waking. This peak is referred to as the cortisol awakening response and is thought to be influenced by light exposure and expected stress levels for the upcoming day (Bernard et al., 2017). This peak is followed by a gradual decline and a dip in the afternoon followed by continued decline into the evening, during which decreased cortisol levels are helpful for increasing tissue repair, decreasing inflammation, and repairing the body for sleep. Afternoon/evening cortisol may be more easily influenced by environmental factors compared to morning cortisol, which is highly hereditary (Van Hulle et al., 2012).
The diurnal cortisol rhythm is an integral part of the circadian rhythm and is believed to develop as early as three months of age and becomes more prominent by preschool age (Clow et al., 2010; Bernard et al., 2017).
In essence, the cortisol system has a basal, rhythmic component that entrains predictability and stability of day-to-day biological processes while the adaptability of the acute stress response allows individuals to respond effectively to environmental changes.
Maltreatment may lead to decreased morning cortisol
The glucocorticoid system is thought to be dysregulated by maltreatment and early adversity. In regard to basal cortisol, three meta-analyses have shown inconsistent effects for adults and children with self-reported maltreatment on plasma (Bernard et al., 2017) or hair cortisol (Khoury et al., 2019) or 24-hour urine sampling (Schär et al., 2022), but in one of the meta-analyses, significant effects emerged with inclusion of 10 studies with confirmed maltreatment histories by a third-party like CPS finding maltreatment was associated with lower wake up cortisol levels in children (g= 0.24; 95% CI: 0.42 to 0.07) but not significantly different cortisol awakening response (CAR) levels or a more flattened diurnal cortisol slope (i.e. decreasing morning cortisol +/- increased evening cortisol) (Bernard et al., 2017). However, one of the other meta-analyses did find higher evening cortisol levels (g = 0.10, 95% CI [0.03; 0.18]), but this effect was no longer significant when removing studies that had included losing a friend or loved one as a form of early adversity (Schär et al., 2022).
Although, one study by the same author suggests that a more flattened diurnal cortisol slope is seen in younger children exposed to maltreatment and that this can be normalized with parental attachment interventions (Bernard et al., 2015). Other studies have also observed flattened diurnal cortisol rhythms, including a recent study of adolescents in which substantiated physical abuse was associated with a more flattened diurnal rhythm (Shirtcliff et al., 2024), as well as studies in monkeys exposed to maternal separation (Sanchez et al., 2005; Feng et al., 2011) There were no observed differences between types of maltreatment in the meta-analysis (abuse vs. neglect).
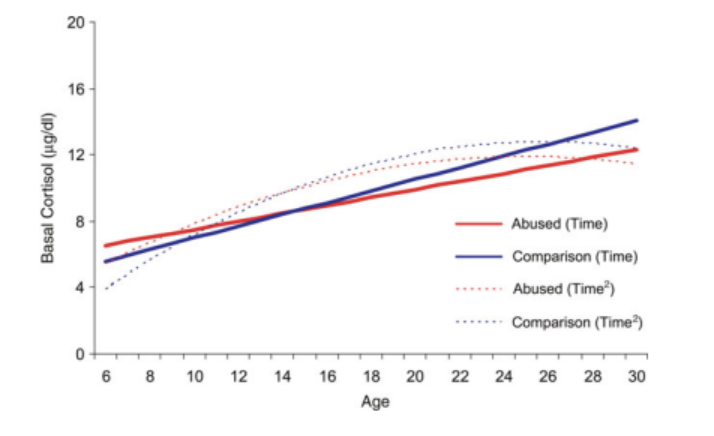
Some studies suggest basal cortisol levels may be higher in children exposed to ACEs before decreasing to subnormal levels in adulthood. One longitudinal study by Tricket et al. (2010) followed sexually abused girls aged 6-16 for 15 years and observed that while basal cortisol levels were initially higher in the sexually abused girls compared to demographically matched controls, these levels showed a significantly slower increase into young adulthood. Normally, basal cortisol increases linearly from childhood to early adulthood before leveling off. This study supports the idea of the attenuation hypothesis, where early hypercortisolism leads to an adaptive downregulation of the glucocorticoid system.
Limitations to these studies
Indeed, whether children exposed to maltreatment experience chronic hyper- or hypocortisolism may depend on the type of maltreatment experienced (abuse vs. neglect) or the proximity that the cortisol was measured in relation to their trauma; but findings have been mixed to date.
Human studies often measure cortisol many years following traumatic exposure. A meta-analysis of trauma-exposed adults showed that with the more months that had elapsed since the stress first emerged, the lower a person’s morning cortisol total cortisol, ACTH to CRH challenge, and post dexamethasone cortisol (Miller et al., 2007). This could confound some studies finding increased cortisol levels in children exposed to ACEs. It's also a sign that the downregulation of the HPA-axis may happen months to years after trauma.
Most human studies oversample females and rely on self-report for maltreatment.
The presence of psychopathology (PTSD, MDD) is often not reported or varies from study to study. These are known moderators of cortisol.
Grouping of participants and assessment of maltreatment severity vary widely from study to study. For instance, some studies may group participants based on whether they had experienced sexual abuse vs. no sexual abuse. The other types of maltreatment can confound this method of grouping, as can unreported maltreatment in studies that rely on third-party reporting.
Maltreatment is associated with blunted cortisol reactivity to psychosocial stress
There have been three recent meta-analyses assessing cortisol reactivity in participants with ACEs.
Bunea et al. (2017) conducted a meta-analysis of 29 studies on cortisol reactivity in children and adults with ACEs (5% with a clinical diagnosis) to the Trier Social Stress Test (TSST), which involves hastily preparing a presentation or speech on a given topic to a panel of “judges” who are instructed to remain neutral. Following the speech, participants often perform challenging arithmetic. The meta-analyses showed that individuals with ACEs have a more blunted cortisol response to psychosocial stressors compared to those without ACE history (g = −0.39). The blunted cortisol response was more prevalent in adults than in children (adults: g = −0.63; 95% CI: –0.97, –0.28) (children: g = −0.25; 95% CI: −0.38, −0.11) and more prevalent in cases of maltreatment (g = −0.49; 95% CI: −0.65, −0.32) than in other ACEs (parental separation, parental mental illness and substance abuse, illness, death of family, etc.) (g = −0.14; 95% CI: −0.24).
A more recent meta-analyses of 35 studies by Schär et al. (2022) corroborated these findings showed a smaller decrease in cortisol reactivity to the TSST, with effect sizes for different measurement parameters (peak cortisol levels, change in cortisol levels, cortisol recovery time, and total cortisol [AUC]) ranging from -0.17 to -0.28. Notably, there were no differences on the dexamethasone suppression test or the dexamethasone-CRH suppression test.
A recent systematic review and meta-analysis grouped studies using the CTQ to measure ACEs, finding greater CTQ scores correlated with a more blunted cortisol reactivity, (k = 29, β = -0.006; 95% CI: −.010 to −.002, p = .01, R2 = .33) (Brindle et al., 2022), suggesting a dose-response relationship.
Although early life stress is theorized to be uniquely impactful on the development of the HPA-axis, there are few human studies supporting this hypothesis. Nevertheless, a longitudinal study by Young et al. (2019) showed that neither cumulative life stress, early life stress alone, nor adult life stress alone predicted blunted cortisol responses to the TSST in adulthood. Current life stress only predicted blunted cortisol response if there was also presence of high early life stress before the age of 5, defined by 1SD above the mean. This suggests early life stress leads to later HPA-axis dysfunction.
Implications of a blunted cortisol response
Rather than being beneficial, a blunted cortisol response can be problematic, indicating an impairment in the body's ability to respond to and recover from stress. This impairment can lead to various long-term health issues, especially if the stressor is chronic or particularly severe. For individuals with a history of adverse childhood experiences (ACEs), this blunted response might reflect a kind of wear and tear on the stress response system, which has been forced to adapt to high levels of stress from an early age.
Similar to individuals with high ACE scores, those with PTSD (d = −.36, SE = .15) (Morris et al., 2012) and BPD (g = -0.32, 95% CI [-0.56 to -0.06]) (Thomas et al., 2018) exhibit reduced basal cortisol levels, and there is a diminished cortisol response in borderline personality disorder to stress (g = -0.32, 95% CI [-0.53, -0.11]) (Thomas et al., 2018) as well as a blunted response to both stress and dexamethasone in PTSD (Morris et al., 2012).
Interestingly, the meta-analyses by Morris et al. (2012) revealed that PTSD patients with comorbid depression exhibit increased cortisol output throughout the day but still experience a pronounced decrease in cortisol in response to dexamethasone. Indeed, different subtypes of depression are associated with different HPA-axis profiles (Gold and Chrousos, 2002; Lamers et al., 2013):
Atypical Depression (mood reactivity, weight gain, hypersomnia, leaden paralysis, sensitivity to rejection) - hypoactive HPA system, lower basal cortisol, response to stress has blunted cortisol response, dexamethasone test more normal suppression
Melancholic Depression (severe anhedonia, early morning awakenings, psychomotor agitation/retardation, significant weight loss, excessive guilt) - hyperactivity/more reactive HPA system, higher basal cortisol, heightened cortisol response, dexamethasone test often shows non-suppression, higher CRH
Aside from its association with increased psychiatric symptoms, a blunted cortisol response has also been associated with increased adiposity, and potentially greater inflammation (Ronaldson et al., 2016).
HPA-Axis Beyond Cortisol: Low Levels Of Cortisol May Increase CRH and Lead To Increased Anxiety, Depression, Fear, And Addictive Behaviors
Most studies of rodents exposed to maternal separation show an increase in lifelong changes to the stress response demonstrated through changes at the molecular and behavioral level. Adult rodents who were exposed to recurrent maternal separation for 3-6 hours during the first two weeks of life show increased depressive behavior, increased anxiety to novelty, and increased/decreased response to stress into adulthood (Korosi and Baram, 2009). One of the notable molecular changes reported in these recurrently separated rodents include increased ACTH and corticosterone responses to acute psychosocial stress or pain, and this effect persists into adulthood but can be rescued with increased physical contact or feeding the pups (Van Oers et al., 1998).
Another potential moderator of the effect of ACEs on the HPA-axis is CRH. Maternal separation in animal models consistently show increased basal levels of CRH in CSF. This may be a result of decreased expression of GRs in both hippocampus and hypothalamus (Plotsky and Meaney, 1993), which may inhibit negative feedback from cortisol, while increasing hypothalamic and amygdala CRH and increasing CRH receptors in the hippocampus, PFC, PVN, locus coeruleus, and raphe nucleus (Korosi and Baram, 2009; Veenema 2009). Similar effects have been observed in monkeys, as increased basal CRH in the cerebrospinal fluid is seen following maternal separation and abuse, despite decreased basal cortisol levels and/or reactivity to stress (Veenema et al., 2009). Multiple other animal models have shown a similar phenotype of blunted basal or reactive cortisol with elevated CRH (Veenema et al., 2009; Orso et al., 2020).
CRH is thought to potentiate anxiety and addictive behaviors through binding to CRF1 receptors in the amygdala, BNST, hippocampus, VTA, as well as the PFC. Although in animal models increased CRH or CRF1 activity has been heavily implicated in depressive, anxiety, and addictive behaviors resulting from early life stress and that many of these behaviors are reversible with CRF1 antagonism, translation to human studies has been limited (Sanders and Nemeroff, 2016).
In 2000s/2010s several phase 2 clinical trials of CRF1 receptor antagonists on depression and anxiety were conducted, with disappointing results. Of the six studies, five had null or negative results and the only trial with positive results was limited by hepatotoxicity (Sanders and Nemeroff, 2016).
Epigenetics And Gene-Environment Interactions: Maltreatment Is Associated With Hypermethylation Of The Glucocorticoid Receptor Gene And Gypomethylation Of FKBP5
The glucocorticoid receptor gene (NR3C1) is hypermethylated in patients with a history of abuse who died by suicide, with reduced expression of glucocorticoid receptors
Rodent studies show increased maternal care behaviors decrease methylation of Nr3c1, leading to increased glucocorticoid expression in the hippocampus and to greater stress tolerance in adulthood
Epigenetic changes may be modifiable, with maltreated preschoolers showing higher NR3C1 methylation initially but normalization over time
FKBP5 is a regulatory protein that increases glucocorticoid receptor resistance
Low levels of FKBP5 are thought to increase risk of developing PTSD and have been associated with worse PTSD symptoms
Carriers of polymorphisms of FKBP5 may be at higher risk of developing PTSD and/or depression in response to early life stress
Studies are mixed as to how FKBP5 alters the HPA-axis
More large-scale studies examining the entire genome are needed to identify genes that increase the risk of disease in response to maltreatment
The epigenetic modifications of genes encoding glucocorticoid receptors (NR3C1) and the proteins that regulate its function (FKBP5) have seen increasing interest in an attempt to explain how early life stress continues to affect the HPA-axis into adulthood.
Methylation is most often associated with decreased gene expression. Animal studies and postmortem human studies report decreased glucocorticoid receptor and mineralocorticoid receptor expression in response to early life stress that may be specific to the hippocampus (Arabadzisz et al., 2010; McGowan et al., 2009; Veenema et al., 2009).

Nuclear Receptor Subfamily 3, Group C, Member 1 gene (NR3C1) From Ibrahim et al., 2012:
Several postmortem studies of patients with abuse history who died by suicide show these patients had reduced expression of glucocorticoid receptors and hypermethylation of NR3C1 promotors in the hippocampus compared to those without abuse history (Takahashi et al., 2018; Ibrahim et al., 2012).
A review of studies involving peripheral blood samples from patients with a history of maltreatment found that 8 out of 11 studies suggested the 1F promoter of NR3C1 is hypermethylated (Wadji et al., 2021), and this finding is consistent across samples of patients with MDD, BPD, PTSD, and without a psychiatric diagnosis.
However, the specific CpG binding sites identified varied between studies, and the overall effect of methylation has not been consistently replicated by epigenome-wide association studies (EWAS) or prospective studies (Houtepen et al., 2018; Marzi et al., 2018; Dunn et al., 2019).
Other studies in BPD patients and patients with adult-onset PTSD have also shown decreased peripheral methylation of NR3C1 promoters (Flasbeck and Brüne, 2021; Yehuda et al., 2015; Schechter et al., 2015, Labonte et al., 2014). Nevertheless, it should be noted that peripheral methylation may not be an adequate proxy for methylation in the brain, as epigenetic modifications likely occur in a tissue-specific, or even cell-specific manner (Lutz and Turecki, 2014).
Rodent studies show that maternal licking grooming of pups increases hippocampal GR and MR expression, increases hippocampal size and function, and decreases adult stress response (Korosi and Baram, 2009) this may be related to decreased methylation and increased acetylation of the 17 promoter (analogous to the 1F promoter in humans) of Nr3c1 in the hippocampus (Weaver et al., 2004; Murgatroyd et al., 2015). This suggests that early life stress, through decreasing GR and MR, may lead to a decreased sensitivity to negative feedback by cortisol.
It should be noted that it’s still uncertain to what extent epigenetic changes are modifiable and how deterministic sensitive periods are. One longitudinal study by Parent et al. (2017) showed that maltreated preschoolers who had experienced documented maltreatment within 6 months of the study showed higher methylation at NR3C1 1D at baseline compared to non-maltreated preschoolers, but levels of methylation decreased to levels equal to or less than the control group 6 months later.
FKBP
There are likely other mechanisms regulating HPA activity aside from altering GR expression in the hippocampus. FKBP5 is believed to decrease sensitivity of the glucocorticoid receptor to glucocorticoids by decreasing the receptor’s ability to bind glucocorticoids and to translocate to the nucleus. FKBP’s expression is also induced by glucocorticoids, meaning it functions in a negative feedback loop with glucocorticoids (Matosin et al., 2018).
Studies have consistently linked lower FKBP5 expression with PTSD risk and have identified low FKBP5 expression as a risk factor for later PTSD development in pre-deployment soldiers even when controlling for childhood trauma (van Zuiden et al., 2012). Post-mortem studies have also found increased FKBP5 expression in the brains of individuals with BPD, MDD, and schizophrenia (Matosin et al., 2018).
A meta-analysis by (Wang et al., 2018) found evidence for an interaction between early adversity and polymorphisms of the FKBP5 gene that increase risk for depression and PTSD. Carriers of different alleles for SNPs (T-rs1360780 [6 studies], G-rs3800373 [4 studies], T-rs9470080 [5 studies], rs9296158 [see Sheerin et al., 2020]) have consistently been shown to increase PTSD risk while only one SNP has been analyzed in depression (T-rs1360780 [8 studies]).
One study by Binder et al. (2008) showed that carriers of the A risk allele for rs9296158, have increased PTSD symptoms in adulthood with increasing childhood trauma but not adulthood trauma. Notably this dose-dependent effect of childhood trauma with the number of A-alleles. Participants with two types of childhood trauma and mean PTSD Symptom Scale (PSS) scores (>20 is considered clinically significant) in adulthood:
No risk allele: 13.54 (SD: 3.76)
Heterozygous carrier of A risk allele: 21.25 (2.03)
Homozygous for A risk allele: 31.11 (5.37)
This interaction of childhood trauma with SNPs could be mediated by epigenetic modifications, as one study observed that higher cortisol levels demethylate FKBP5 intron 7 in carriers of the SNP rs1360780 T risk allele and that higher CTQ scores, but not adulthood trauma, were correlated with lower methylation (Klengel and Binder, 2014). Although, this study was relatively smaller (N=76).
FKBP5 may also have implications for psychotherapeutic outcomes in PTSD patients, as Levy-Gigi et al. (2013) noted increased FKBP5 expression and hippocampal size as predictors of treatment response in PTSD patients following 12 weekly 1.5-hour sessions of trauma-focused cognitive behavioral therapy. Likewise, in a study of 43 Ugandans with PTSD undergoing narrative exposure therapy, carriers of the FKBP5 rs1360780 T-risk allele experienced significantly less improvement in PTSD symptoms compared to noncarriers at 10-month post treatment (Cohen's D=1.23 vs. 3.72) (Wilker et al., 2014).
Although FKBP5 has been implicated in increasing risk for depression and PTSD, the mechanism underlying this increased risk is not yet clear. Klengel and Binder showed that the T-risk allele of rs1360780 allele alters the interaction of transcription factors at the promoter of FKBP5, leading to increased transcription of FKBP5 (Klengel and Binder, 2014) and other groups have shown the four risk alleles increase FKBP5 expression in response to glucocorticoids (Matosin et al., 2018). The overexpression of FKBP5 is theorized to delay the negative feedback of glucocorticoids on the HPA-axis through increasing glucocorticoid resistance, which would lead to a prolonged increase in HPA-axis activity.
Meanwhile, Sarapas et al. (2011) observed homozygosity for any one of the four risk alleles was associated decreased FKBP5 expression (B = −177.91), which in turn was correlated with decreased cortisol (B = 0.01) , and increased PTSD symptoms (B = −0.03). These findings are in alignment with studies in PTSD patients showing decreased FKBP5 expression is a risk factor for worse outcomes.
Some authors note that while these SNPs cause glucocorticoid resistance in healthy individuals through FKBP5 overexpression, they are also linked to increased sensitivity to glucocorticoids in PTSD patients, as seen in dexamethasone suppression tests (Vale and Carvalho, 2020). This indicates that other factors influence FKBP5 functionality or that additional factors contribute to the HPA-axis phenotype observed in PTSD patients.
It should be noted that studies looking at one or two genes and their epigenetic modifications–also known as candidate-gene studies–have seen a high degree of criticism in recent years due to their risk for producing false positive results. This is due to their high reliance on regression statistics and inability to control for many important covariates. This leads to a high amount of publication bias. Trauma, increased aerobic exercise, dietary changes, and other physiologic insults produce widespread changes in thousands of epigenetic markers, making it unlikely that variants in one or two genes produce the majority of an effect. Thus, more studies need to assess the effects of ACEs on the entire genome through an epigenome wide approach (EWAS). There have been several EWASes looking at the effects of ACEs, and they do not find the same associations in candidate gene studies of NR3C1 or FKBP5, and have yet to find any consistent association with ACEs (Houtepen et al., 2018; Marzi et al., 2018; Dunn et al., 2019). However, they may currently be underpowered to detect subtle, consistent changes in the epigenome, and more studies in the future will need larger sample sizes. Nevertheless, given the theorized functional relevance of FKBP5 in modulating the glucocorticoid receptor’s response to cortisol, functional polymorphisms in the gene likely do have some effect on the HPA axis, albeit a smaller one than is currently attributed and one that is difficult to measure.

ACEs Are Associated With Decreased Hippocampal Volume
The amygdala, hippocampus, and prefrontal cortices are the regions with the highest expression of glucocorticoid receptors and are some of the most highly studied brain regions in relation to chronic stress and neurodevelopment. The hippocampus specifically has been shown to reduce in size in response to chronic glucocorticoid and/or CRH stimulation in animal models (Sapolsky, 1996, Maras and Baram, 2012).
According to a 2022 review of meta-analyses by Hakamata et al. 2022, there have been 7 meta-analyses assessing total brain gray matter volume changes in adults with ACEs history compared to those without.
All seven have shown decreased hippocampal volumes in the group with ACEs, with a small effect size averaging 0.25 (range from .08 to .77). Findings for the amygdala and other brain regions, including PFC regions, were inconsistent, but more studies tended to report reductions.
One of these seven meta-analyses (Calem et al., 2017) noted a small effect size of ACEs and decreased hippocampal volume when including 9 studies with 1200 participants without any past psychiatric history (g =- 0.15 [95 CI -0.26 to -0.03]). But this became insignificant when controlling for gender (g = −0.06, p = 0.368)
It should be noted that most of these meta-analytic studies did not assess dose-response, and that the majority of studies included participants who experienced a single trauma. Riem et al. (2015) was the only meta-analysis that looked at the role of multiple trauma exposure compared to single trauma exposure. They assessed only hippocampal volume changes, and despite the fact this study reported low effect sizes (.09), they found that only participants with multiple exposure had reduction in hippocampal volume while those with single exposure did not. Thus, more studies assessing dose-response are needed.
These studies also suggest that children and adolescents do not have lower hippocampal volumes, but rather the decrease in volume is observed into adulthood, which is consistent with the idea that neurogenesis continues in the hippocampus throughout early adulthood.

What is the significance of a .25 SD difference in hippocampal volume?
According to a series of meta-analyses by (Schmaal et al., 2016; Ho et al., 2022), patients with MDD (d=−0.14, % difference=−1.24), recurrent MDD (d=−0.17, % difference=−1.44), and early-onset MDD (d=−0.20, % difference=−1.85) have smaller hippocampal volumes, specifically in CA1 regions, which are known to be responsive to stress and are important for learning and memory.
A meta-analysis by Nelson and Tumpap (2017) showed that PTSD patients have decreased hippocampal volume (left hippocampus effect size=–0.400, p<0.001, 5.24% reduction; right hippocampus effect size=–0.462, p<0.001, 5.23% reduction) and that Caps-5 scores correlated negatively with left hippocampal volume decreases but not the right.
There is debate that decreased hippocampal volume observed in PTSD and depression is a result of early trauma and not intrinsic to these disorders. Teicher's endophenotype concept suggests that individuals with a history of maltreatment represent unique biological subtypes of depression, distinct from those with depression but no maltreatment history. He argues that the significant brain differences observed in depressed patients, such as decreased hippocampal volume, might be confounded by the effects of maltreatment rather than being intrinsic markers of depression itself (Teicher and Samson 2013).
One theory of how reduced hippocampal volume could lead to psychopathology is through the lens of reduced pattern distinction. The premise of this theory is that the hippocampus’s primary function is in pattern distinction, which is vital to learning and memory. If this function is decreased, and in the setting of a bias towards negative stimuli, which is commonly seen in patients with depression, this could lead to overgeneralization of ambiguous social or environmental cues towards a negative perception (Lecei and van Winkel, 2020). If negative stimuli are more consistently encoded and neutral/positive stimuli are not, this could subsequently lead to persistent negative narratives about the self and the world.
Indeed, several fMRI studies report that participants with high CTQ scores display differential activity in the left hippocampus in response to novel stimuli, whether it be new faces or odd-ball tasks (Edmiston et al., 2013; Derome et al., 2023).
Exercise and environmental enrichment increase hippocampal volume
Numerous cross-sectional, observational, and RCTs in animals and humans have shown exercise increases gray matter volume in the hippocampus (Erickson et al. 2014).
For example, one RCT by Erickson et al. (2011) of older adults showed that brisk walking for one year not only attenuated age-related gray matter decline as observed in the stretching group, but that it led to a 1-2% increase in hippocampal volume. These increases also correlated to greater spatial memory.
There are also animal models supporting the notion that environmental enrichment, which involves providing a stimulating environment with opportunities for physical activity, social interactions, and cognitive challenges, could lead to increased hippocampal volume and synaptic plasticity (Olson et al., 2006; Eckert and Abraham, 2013).
IQ was not changed by trauma when controlled for family socioeconomic status (Danese et al., 2016) except in cases of severe neglect (Perry 2002)
A prospective study by Danese et al. (2016) followed two different cohorts, one in the UK (N=2234) and one in New Zealand (N=1037), to assess the effect of maltreatment on IQ and cognitive performance into adulthood. In the U.K. twin cohort, children who had experienced documented poly-victimization between 5-12 years old had lower IQ scores at age 12 compared to those without victimization (beta = −0.17, p<0.01), but this effect was attenuated after controlling for IQ at age 5 and family SES (beta = −0.05, p = 0.02). This trend held true for IQ at age 18, as well as performance on multiple cognitive tasks on the Cambridge Neuropsychological Test Automated Battery including spatial working memory, spatial span, and processing latency, where IQ at age 5 and family SES were significant predictors (betas averaging .2 to .3).
This trend was again observed in the New Zealand cohort, finding the effects of childhood trauma on IQ at age 38, working memory, processing speed, verbal comprehension, and perceptual reasoning became insignificant when controlling for maternal IQ at age 3 and family SES.
These effects held true for both prospective and retrospective (CTQ) reporting.
Developmental Outcomes in Romanian Adoptees after being adopted:

Figure 1 above comes from Perry 2002 showing dramatic differences of neglect.
The study by Rutter et al, 1998, "Developmental Catch-up and Deficit Following Adoption after Severe Global Early Privation" involved two groups of Romanian children adopted into the UK, categorized based on their age at the time of adoption. The conditions in the residential institutions labeled as "hospitals" or "orphanages" were devastatingly horrific. In these facilities, children, abandoned for various reasons, were largely confined to cots, devoid of toys or playthings, and experienced minimal interaction or speech from caregivers. Caregiving lacked any form of personalization, with basic needs like feeding often reduced to gruel served in bottles with large teats, sometimes left propped up for the children to feed themselves. The physical environment was often harsh, with practices such as washing the children by hosing them down with cold water, further illustrating the neglect and severe lack of humane care in these institutions.
Upon their arrival in the U.K., the majority of the children were in a deteriorated physical condition. They suffered from severe malnutrition, skin disorders, respiratory infections, and chronic intestinal infections, such as giardiasis. They had around 2 standard deviations lower weight, height and head circumference compared to English children. Their Denver quotient for those adopted before age 2 was 76.5, and those adopted between 24 and 42 months was 48.1.
Retesting at 4 years showed the impact of the enriched environments and the reversal of the impact of their early horrific conditions. While they had almost normal weight and height, their head circumference remained around 1-1.5 SD below the mean (worse if entry was after 6 months). The IQ increase was profound and breaths hope into the impact of an enriched environment. The Denver quotient was 115.7 for those who came before 6 months and 96.7 for those that came after 6 months.
Adults who have experienced extreme childhood abuse and neglect can also show immense response to the supportive environments at partial hospitalization programs. For example, Bateman et al. (2008) (see episode 206) showed that 432 hours of therapy (72hr of individual mentalization based therapy [MBT], 216hr of group MBT, 144 hr of expressive therapy and community meeting) over 1.5 years and an optional booster of 2 hours/week (144 hr) of group MBT for 18 months lead to significant improvements in BPD symptoms (d =1.80), suicide attempts (d = 1.4), hospitalizations (d = 1.50), employment (d = 0.94), and decreased use of antidepressants (d =1.10), antipsychotics (d = 2.04), and mood stabilizers (d = 1.17) at assessment five years after discharge.
Although 600 hours of therapy might seem extensive, it is minimal compared to the countless hours patients endured abuse and neglect during childhood. As discussed in episode 206, these 600 hours of therapy become even more impactful when considering patients can apply these mentalization-based skills to improving their relationships outside of therapy. Thousands of hours spent in deeper connection with friends and family is therapeutic in and of itself and can heal the damages of early life trauma.
ACEs Decrease White Matter Integrity In The Corpus Callosum
White matter is the long collection of myelinated axons that connect different parts of the brain over long distances, such as between the two hemispheres. The corpus callosum is the largest white matter tract in the brain and is important for interhemispheric communication between cortices. Lim et al. (2020) conducted a meta-analysis of whole-brain diffusion tensor imaging (DTI) studies (or studies that look at the diffusion of water through the brain) and showed that those with ACEs had significant decreases of fractional anisotropy in several brain regions including:
Corpus callosum
Bilateral thalamus and fornix
Bilateral optic radiations
They also showed that these white matter changes in the corpus callosum may be found in participants without psychiatric comorbidities, according to an underpowered exploratory analysis of three studies.
Interestingly, age was negatively correlated with FA in the bilateral occipital clusters, right thalamus-fornix, and callosal genu and body, and these abnormalities were noted in older but not younger maltreated individuals compared to age-matched controls.
One longitudinal study by Goetschius et al., 2020 “Childhood violence exposure and social deprivation predict adolescent amygdala-orbitofrontal cortex white matter connectivity” looked at white matter differences in 183 adolescents (15–17 years) whose mothers were interviewed at the child’s birth and again when the child was 1, 3, 5, 9, and 15 years of age on the child’s experience of violence exposure and social deprivation. Using diffusion MRI technology, a modality with greater sensitivity than DTI at detecting subtle changes in white matter morphology, the study showed that when adjusting for recent life stress, internalizing psychopathology, gender, race, maternal education at birth, and maternal marital status at birth, violence exposure and social deprivation at ages 3, 5, and 9 significantly predicted the probability of right hemisphere amygdala–OFC white matter connectivity at age 15-17 (β =−0.317, p = 0.005).
When social deprivation was 0.78 standard deviations or greater, violence exposure and degree of white matter connectivity were inversely related (β = −0.29, p = 0.048). This means that kids with a lack of community or home emotional support, witnessing or being a victim of home or community violence led to less development of white matter connections between amygdala and orbitofrontal cortex, and therefore, greater activation of amygdala to threatening faces.
When social deprivation was 1 standard deviation below the mean, there was no association between violence exposure and amygdala–OFC white matter connectivity (β = 0.02, p = 0.209), suggesting social support may act as a buffer for the development of this white matter tract, which was shown to be predictive of amygdala activation to threatening faces in this study, such that increased white matter was correlated with decreased amygdala activation to threatening (fearful and angry) faces (β = −0.290).
ACEs may affect the white matter by directly impacting oligodendrocytes, as rodent models of social isolation during adolescence demonstrated reduced branching of oligodendrocytes (Makidonen et al., 2012), and postmortem studies show differential transcription activity in oligodendrocytes of patients with ACEs who died by suicide, reflecting a more mature phenotype (Tanti et al., 2018).
ACEs Increase Chronic Inflammatory Cytokines
It’s hypothesized that the neuroimmune system is highly influential in shaping myelination and the integrity of white matter tracts, as supported in animal models (Favrais et al., 2011). Glial cells, for example, are thought to directly contribute to neuronal differentiation, neurogenesis, neuroplasticity, synaptic pruning, myelination, and neural metabolism (Araque and Navarrete, 2010). Disruption to these processes, especially in times of neural development, could theoretically lead to long standing changes (Danese and Baldwin, 2017).
Baumeister et al. (2016) conducted a meta-analysis of 18 cross-sectional studies (N=16 870 adults) for C-reactive protein (CRP), 15 studies (N=3751) for interleukin-6 (IL-6), and 10 studies (N=881) tumor necrosis factor-α (TNF-α). Controlling for age, BMI, and gender, their results showed small increases in baseline peripheral levels of these cytokines (CRP: Fisher’s z=0.10, CI [0.05–0.14]; IL-6: z=0.08, CI [0.03–0.14]; TNF-α z=0.23, CI [0.14–0.32]). This meta-analysis also suggested that different ACEs may predispose to increases in different cytokines, with abuse being associated with increased TNF-α and IL-6, but not CRP, while CRP was elevated in parental neglect.
Later, Kuhlman et al. (2020) corroborated some of these findings by conducting a meta-analysis of 12 studies measuring inflammatory markers in children (9 for CRP and 4 for Il-6). Findings revealed a small association for early life adversity and increased CRP (z = .07 [.04, .10]), but not IL-6 (z = .17 [−.07, .42]).
These meta-analyses have since been followed up by several prospective cohort studies in children showing mixed findings and small associations (Reid et al. 2020; Slopen et al., 2012; Priest et al., 2022), with some prospective studies suggesting that the effects of ACEs may be attenuated over time as the individual ages. Meanwhile others suggest that those with exposure to neglect or 4+ ACEs age at a slightly faster rate (Mian et al., 2022). There is also evidence that people with ACEs are more likely to suffer from cognitive decline and dementia (Corney et al., 2022).

It should be noted that many of these studies do not account for confounders related to sleep, smoking, diet, exercise, or socioeconomic status. Inflammation has also not yet been linked to disrupted neural development in humans.
BDNF has also been studied, and most studies have shown no differences plasma/blood levels in those with high maltreatment history vs low/no maltreatment history (Vyas et al., 2023), while some authors argue for a mediating role of decreased BDNF in the relationship between maltreatment and psychopathology in adulthood (Watt et al., 2020; Maes et al., 2023).
ACEs Inconsistently Impact FMRI Resting-State Connectivity Of Large-Scale Brain Networks
As far as our understanding goes for large-scale network connectivity (default mode network [mPFC, PCC, temporoparietal junction], salience network [insular cortex, ACC, amygdala], central executive/frontoparietal network [dlPFC, posterior parietal cortex]) there is very limited understanding for how these networks normally develop in children, let alone in response to trauma (Holz et al., 2023).
Conventionally, depression is associated with increased intrinsic connectivity of the DMN, which may be related to rumination (Hamilton et al., 2015) and decreased intrinsic connectivity of the CEN/FPN (Liston et al., 2009; Kaiser et al., 2015), which may account for cognitive dysfunction.
Meanwhile, PTSD and BPD are thought to be characterized by decreased resting state connectivity within anterior and posterior parts of the DMN, which may indicate altered self-referential processing or recall of autobiographical memories, and increased salience network activity that may be related to hyperarousal (Koch et al., 2016; Shafie et al., 2022). There is inconsistent evidence detailing the functional changes in these networks in response to maltreatment specifically (for review see: Ross et al., 2021), but changes in connectivity between anterior and posterior DMN have also been observed in healthy adults with ACEs (Phillip et al., 2012) and in women with PTSD resulting from sexual abuse in childhood (Bluhm et al., 2008).
More consistent evidence exists for task-induced changes in corticolimbic connectivity (i.e. PFC with hippocampus and amygdala) that may be mediated by increased inflammation (Kraynak et al., 2019), but the directionality of these changes varies with age (Holz et al., 2023).
ACEs Increase Amygdala Reactivity To Negative Emotional Stimuli
Four meta-analytic studies have compared those with ACE history and those without on brain activation during task-based fMRI (Mothersill and Donohoe, 2016; Heany et al., 2018; Hein and Monk, 2017; Kraaijenvanger et al., 2020). All studies included were cross-sectional and retrospectively measured ACEs. These studies were primarily analyses of passive emotional tasks, most commonly viewing emotional faces, although there was a high degree of heterogeneity in the methods used.
Results have primarily converged on the amygdala, with all meta-analytic studies supporting increased activation in participants with a history of ACEs, especially during negative stimuli. Effect sizes were not calculated due to heterogeneity.
2/4 meta-analyses distinguished healthy volunteers from patients. Both meta-analyses showed persistent amygdala hyperactivation. However, it was not clear how “healthy” participant was defined.
One meta-analysis (Heany et al., 2018) specifically looked at studies using CTQ and socioaffective cues (faces, voices, pain)
Results revealed those with high CTQ compared to low CTQ showed:
Increased activation in the left superior frontal gyrus and left middle temporal gyrus
Decreased activation in the left hippocampus and left superior parietal lobule
Right amygdala and left ventral ACC activation was positively related to CTQ scores in response to negative faces
Heightened right amygdala reactivity in response to negative/neutral psychosocial cues have also been observed in patients with BPD and PTSD (Schulze et al., 2018)
Some authors theorize that because heightened amygdala activity is a risk factor for later depression, it represents a tendency towards a more negative perceptual bias (Mattson et al., 2017). Indeed, children exposed to violence tend to have a greater sensitivity to identifying anger and classify more faces displaying negative emotions as angry and attribute more hostile intentions, and this may be mediated by heightened amygdala reactivity (Dodge et al., 1990; Dodge et al., 1995).
ACEs Decrease Reward Anticipation
There have been seven studies assessing reward system activity via fMRI and the monetary incentive task. The standard version of the monetary incentive task involves prompting the participant to hit a button after seeing a flash of light. If the participant is fast enough they will receive a reward. Before the light flash is presented, the participant is cued about the degree of value for the reward. This allows researchers to study the anticipatory aspects of attaining a reward and the effects of receiving the reward.
Six of the seven studies on adults and adolescents with early childhood family adversity (Boeker et al., 2014; Holz et al., 2017; Hanson et al., 2015), emotional neglect (Hanson et al., 2015), or institutionalization (Goff et al., 2013; Mehta et al., 2010) have shown reduced activity of the ventral striatum during the anticipation of a reward while one study showed increased activity (Dennison et al. 2017). Only one study reported a beta of −0.26 for level of adversity and ventral striatum activation, and most studies reported r values around -.2 to -.3 (Hanson et al., 2015). This study also suggested that life stress experienced in early childhood (ages 5-8) was most highly correlated with blunted ventral striatum activity (β−0.327, P = 0.009), while stress experienced in late childhood (β = 0.015, P = 0.9) and adolescence (β = 0.08, P = 0.57) were not statistically correlated.
These results are interesting given decreased striatal activation during reward anticipation is seen in people with substance use disorder (Luijten et al., 2017), anhedonia (Pizzagalli, 2022), and with inflammatory stressors (Eisenberger et al., 2010). Also, considering the striatum receives a high amount of unidirectional input from the amygdala and PFC and that these brain regions all undergo protracted development into adolescence, it seems plausible that ACEs disrupt the development of these circuits.
One potential contributing mechanism to this perturbation could be through modulation of endogenous opioid receptors.
Evidence primarily from animal models suggests that maternal separation modulates the expression and sensitivity of endogenous opioid receptors, leading to increased pain sensitivity and addictive behaviors (Nakamoto and Tokuyama, 2023). In humans, postmortem biopsies of depressed patients who committed suicide and had a history of childhood abuse showed decreased expression of kappa opioid receptors in the anterior insula compared to those without a history of abuse (Lutz et al., 2018), and one study comparing the subjective effects of morphine suggested that healthy participants with a high CTQ score reported liking the drug more than those with low CTQ scores (Carlyle et al., 2021).
These biological changes in those with ACEs may explain some of their propensity for developing a substance use disorder later in life (see episode 204) and should increase our compassion for those with substance use disorders. Decreasing the shame felt by patients who struggle with addiction is vital for promoting connection, as it is a chronic relapsing disorder. Cultivating a stable, longitudinal connection with these patients and increasing their mentalizing capabilities can help them to overcome addiction through enhancing their protective relationships outside of therapy.
Conclusion
ACEs undeniably leave a lasting impact on a person's biology. However, the effects on the HPA-axis or the hippocampus may not be the most crucial aspects of ACEs' impact on the brain. ACEs likely alter cortical development in ways that make abilities like mentalizing and self-regulation more challenging. These capabilities are difficult to measure through neuroimaging but are vital for fostering deep, intimate connections. As a result, patients who have experienced early life traumas often struggle with forming and maintaining meaningful relationships, impacting their ability to connect with others throughout their lives. It is these abilities that we should strive to improve in our patients as mental health professionals. Improving our patients' capacity to reflect on the mental states and intentions of attachment figures and others, can have profound impacts on their relationships outside of therapy. This progress is crucial for reversing the impacts of childhood trauma.
As mental health professionals, we have a responsibility to facilitate these improvements. Our ability to do so depends not only on our clinical skills but also on our capacity to manage our own emotional responses and be patient and fully present with our patients, especially in the face of difficult interpersonal situations that inevitably arise in therapy. By managing our countertransference, addressing vicarious trauma, and improving our reflective functioning through engaging in our own therapy, we can create a therapeutic environment that promotes the healing and growth of our patients.
References
Arabadzisz, D., Diaz-Heijtz, R., Knuesel, I., Weber, E., Pilloud, S., Dettling, A. C., Feldon, J., Law, A. J., Harrison, P. J., & Pryce, C. R. (2010). Primate early life stress leads to long-term mild hippocampal decreases in corticosteroid receptor expression. Biological psychiatry, 67(11), 1106–1109. https://doi.org/10.1016/j.biopsych.2009.12.016
Araque, A., & Navarrete, M. (2010). Glial cells in neuronal network function. Philosophical transactions of the Royal Society of London. Series B, Biological sciences, 365(1551), 2375–2381. https://doi.org/10.1098/rstb.2009.0313
Bateman, A., & Fonagy, P. (2008). 8-year follow-up of patients treated for borderline personality disorder: mentalization-based treatment versus treatment as usual. The American journal of psychiatry, 165(5), 631–638. https://doi.org/10.1176/appi.ajp.2007.07040636
Baumeister, D., Akhtar, R., Ciufolini, S., Pariante, C. M., & Mondelli, V. (2016). Childhood trauma and adulthood inflammation: a meta-analysis of peripheral C-reactive protein, interleukin-6 and tumour necrosis factor-α. Molecular psychiatry, 21(5), 642–649. https://doi.org/10.1038/mp.2015.67
Bernard, K., Frost, A., Bennett, C. B., & Lindhiem, O. (2017). Maltreatment and diurnal cortisol regulation: A meta-analysis. Psychoneuroendocrinology, 78, 57–67. https://doi.org/10.1016/j.psyneuen.2017.01.005
Bernard, K., Hostinar, C. E., & Dozier, M. (2015). Intervention effects on diurnal cortisol rhythms of Child Protective Services-referred infants in early childhood: preschool follow-up results of a randomized clinical trial. JAMA pediatrics, 169(2), 112–119. https://doi.org/10.1001/jamapediatrics.2014.2369
Binder, E. B., Bradley, R. G., Liu, W., Epstein, M. P., Deveau, T. C., Mercer, K. B., Tang, Y., Gillespie, C. F., Heim, C. M., Nemeroff, C. B., Schwartz, A. C., Cubells, J. F., & Ressler, K. J. (2008). Association of FKBP5 polymorphisms and childhood abuse with risk of posttraumatic stress disorder symptoms in adults. JAMA, 299(11), 1291–1305. https://doi.org/10.1001/jama.299.11.1291
Bluhm, R. L., Williamson, P. C., Osuch, E. A., Frewen, P. A., Stevens, T. K., Boksman, K., Neufeld, R. W., Théberge, J., & Lanius, R. A. (2009). Alterations in default network connectivity in posttraumatic stress disorder related to early-life trauma. Journal of psychiatry & neuroscience : JPN, 34(3), 187–194.
Boecker, R., Holz, N. E., Buchmann, A. F., Blomeyer, D., Plichta, M. M., Wolf, I., Baumeister, S., Meyer-Lindenberg, A., Banaschewski, T., Brandeis, D., & Laucht, M. (2014). Impact of early life adversity on reward processing in young adults: EEG-fMRI results from a prospective study over 25 years. PloS one, 9(8), e104185. https://doi.org/10.1371/journal.pone.0104185
Brindle, R. C., Pearson, A., & Ginty, A. T. (2022). Adverse childhood experiences (ACEs) relate to blunted cardiovascular and cortisol reactivity to acute laboratory stress: A systematic review and meta-analysis. Neuroscience and biobehavioral reviews, 134, 104530. https://doi.org/10.1016/j.neubiorev.2022.104530
Bunea, I. M., Szentágotai-Tătar, A., & Miu, A. C. (2017). Early-life adversity and cortisol response to social stress: a meta-analysis. Translational psychiatry, 7(12), 1274. https://doi.org/10.1038/s41398-017-0032-3
Calem, M., Bromis, K., McGuire, P., Morgan, C., & Kempton, M. J. (2017). Meta-analysis of associations between childhood adversity and hippocampus and amygdala volume in non-clinical and general population samples. NeuroImage. Clinical, 14, 471–479. https://doi.org/10.1016/j.nicl.2017.02.016
Carlyle, M., Broomby, R., Simpson, G., Hannon, R., Fawaz, L., Mollaahmetoglu, O. M., Drain, J., Mostazir, M., & Morgan, C. J. A. (2021). A randomised, double-blind study investigating the relationship between early childhood trauma and the rewarding effects of morphine. Addiction biology, 26(6), e13047. https://doi.org/10.1111/adb.13047
Castro-Vale, I., & Carvalho, D. (2020). The Pathways between Cortisol-Related Regulation Genes and PTSD Psychotherapy. Healthcare (Basel, Switzerland), 8(4), 376. https://doi.org/10.3390/healthcare8040376
Clow, A., Hucklebridge, F., Stalder, T., Evans, P., & Thorn, L. (2010). The cortisol awakening response: more than a measure of HPA axis function. Neuroscience and biobehavioral reviews, 35(1), 97–103. https://doi.org/10.1016/j.neubiorev.2009.12.011
Corney, K. B., West, E. C., Quirk, S. E., Pasco, J. A., Stuart, A. L., Manavi, B. A., Kavanagh, B. E., & Williams, L. J. (2022). The Relationship Between Adverse Childhood Experiences and Alzheimer's Disease: A Systematic Review. Frontiers in aging neuroscience, 14, 831378. https://doi.org/10.3389/fnagi.2022.831378
Danese, A., & Baldwin, J. R. (2017). Hidden Wounds? Inflammatory Links Between Childhood Trauma and Psychopathology. Annual review of psychology, 68, 517–544. https://doi.org/10.1146/annurev-psych-010416-044208
Danese, A., Moffitt, T. E., Arseneault, L., Bleiberg, B. A., Dinardo, P. B., Gandelman, S. B., Houts, R., Ambler, A., Fisher, H. L., Poulton, R., & Caspi, A. (2017). The Origins of Cognitive Deficits in Victimized Children: Implications for Neuroscientists and Clinicians. The American journal of psychiatry, 174(4), 349–361. https://doi.org/10.1176/appi.ajp.2016.16030333
Dennison, M. J., Sheridan, M. A., Busso, D. S., Jenness, J. L., Peverill, M., Rosen, M. L., & McLaughlin, K. A. (2016). Neurobehavioral markers of resilience to depression amongst adolescents exposed to child abuse. Journal of abnormal psychology, 125(8), 1201–1212. https://doi.org/10.1037/abn0000215
Derome, M., Machon, S., Barker, H., Kozhuharova, P., Orlov, N., Morgenroth, E., Hugdahl, K., & Allen, P. (2023). High levels of childhood trauma associated with changes in hippocampal functional activity and connectivity in young adults during novelty salience. European archives of psychiatry and clinical neuroscience, 273(5), 1061–1072. https://doi.org/10.1007/s00406-023-01564-3
Dodge, K. A., Bates, J. E., & Pettit, G. S. (1990). Mechanisms in the cycle of violence. Science (New York, N.Y.), 250(4988), 1678–1683. https://doi.org/10.1126/science.2270481
Dodge, K. A., Pettit, G. S., Bates, J. E., & Valente, E. (1995). Social information-processing patterns partially mediate the effect of early physical abuse on later conduct problems. Journal of abnormal psychology, 104(4), 632–643. https://doi.org/10.1037//0021-843x.104.4.632
Dunn, E. C., Soare, T. W., Zhu, Y., Simpkin, A. J., Suderman, M. J., Klengel, T., Smith, A. D. A. C., Ressler, K. J., & Relton, C. L. (2019). Sensitive Periods for the Effect of Childhood Adversity on DNA Methylation: Results From a Prospective, Longitudinal Study. Biological psychiatry, 85(10), 838–849. https://doi.org/10.1016/j.biopsych.2018.12.023
Edmiston, E. K., & Blackford, J. U. (2013). Childhood maltreatment and response to novel face stimuli presented during functional magnetic resonance imaging in adults. Psychiatry research, 212(1), 36–42. https://doi.org/10.1016/j.pscychresns.2012.11.009
Eckert, M. J., & Abraham, W. C. (2013). Effects of environmental enrichment exposure on synaptic transmission and plasticity in the hippocampus. Current topics in behavioral neurosciences, 15, 165–187. https://doi.org/10.1007/7854_2012_215
Eisenberger, N. I., Berkman, E. T., Inagaki, T. K., Rameson, L. T., Mashal, N. M., & Irwin, M. R. (2010). Inflammation-induced anhedonia: endotoxin reduces ventral striatum responses to reward. Biological psychiatry, 68(8), 748–754. https://doi.org/10.1016/j.biopsych.2010.06.010
Erickson, K. I., Leckie, R. L., & Weinstein, A. M. (2014). Physical activity, fitness, and gray matter volume. Neurobiology of aging, 35 Suppl 2, S20–S28. https://doi.org/10.1016/j.neurobiolaging.2014.03.034
Erickson, K. I., Voss, M. W., Prakash, R. S., Basak, C., Szabo, A., Chaddock, L., Kim, J. S., Heo, S., Alves, H., White, S. M., Wojcicki, T. R., Mailey, E., Vieira, V. J., Martin, S. A., Pence, B. D., Woods, J. A., McAuley, E., & Kramer, A. F. (2011). Exercise training increases size of hippocampus and improves memory. Proceedings of the National Academy of Sciences of the United States of America, 108(7), 3017–3022. https://doi.org/10.1073/pnas.1015950108
Favrais, G., van de Looij, Y., Fleiss, B., Ramanantsoa, N., Bonnin, P., Stoltenburg-Didinger, G., Lacaud, A., Saliba, E., Dammann, O., Gallego, J., Sizonenko, S., Hagberg, H., Lelièvre, V., & Gressens, P. (2011). Systemic inflammation disrupts the developmental program of white matter. Annals of neurology, 70(4), 550–565. https://doi.org/10.1002/ana.22489
Feng, X., Wang, L., Yang, S., Qin, D., Wang, J., Li, C., Lv, L., Ma, Y., & Hu, X. (2011). Maternal separation produces lasting changes in cortisol and behavior in rhesus monkeys. Proceedings of the National Academy of Sciences of the United States of America, 108(34), 14312–14317. https://doi.org/10.1073/pnas.1010943108
Flasbeck, V., & Brüne, M. (2021). Association between childhood maltreatment, psychopathology and DNA methylation of genes involved in stress regulation: Evidence from a study in Borderline Personality Disorder. PloS one, 16(3), e0248514. https://doi.org/10.1371/journal.pone.0248514
Goetschius, L. G., Hein, T. C., Mitchell, C., Lopez-Duran, N. L., McLoyd, V. C., Brooks-Gunn, J., McLanahan, S. S., Hyde, L. W., & Monk, C. S. (2020). Childhood violence exposure and social deprivation predict adolescent amygdala-orbitofrontal cortex white matter connectivity. Developmental cognitive neuroscience, 45, 100849. https://doi.org/10.1016/j.dcn.2020.100849
Goff, B., Gee, D. G., Telzer, E. H., Humphreys, K. L., Gabard-Durnam, L., Flannery, J., & Tottenham, N. (2013). Reduced nucleus accumbens reactivity and adolescent depression following early-life stress. Neuroscience, 249, 129–138. https://doi.org/10.1016/j.neuroscience.2012.12.010
Gold, P. W., & Chrousos, G. P. (2002). Organization of the stress system and its dysregulation in melancholic and atypical depression: high vs low CRH/NE states. Molecular psychiatry, 7(3), 254–275. https://doi.org/10.1038/sj.mp.4001032
Hakamata, Y., Suzuki, Y., Kobashikawa, H., & Hori, H. (2022). Neurobiology of early life adversity: A systematic review of meta-analyses towards an integrative account of its neurobiological trajectories to mental disorders. Frontiers in neuroendocrinology, 65, 100994. https://doi.org/10.1016/j.yfrne.2022.100994
Hamilton, J. P., Farmer, M., Fogelman, P., & Gotlib, I. H. (2015). Depressive Rumination, the Default-Mode Network, and the Dark Matter of Clinical Neuroscience. Biological psychiatry, 78(4), 224–230. https://doi.org/10.1016/j.biopsych.2015.02.020
Hanson, J. L., Albert, D., Iselin, A. M., Carré, J. M., Dodge, K. A., & Hariri, A. R. (2016). Cumulative stress in childhood is associated with blunted reward-related brain activity in adulthood. Social cognitive and affective neuroscience, 11(3), 405–412. https://doi.org/10.1093/scan/nsv124
Hanson, J. L., Hariri, A. R., & Williamson, D. E. (2015). Blunted Ventral Striatum Development in Adolescence Reflects Emotional Neglect and Predicts Depressive Symptoms. Biological psychiatry, 78(9), 598–605. https://doi.org/10.1016/j.biopsych.2015.05.010
Heany, S. J., Groenewold, N. A., Uhlmann, A., Dalvie, S., Stein, D. J., & Brooks, S. J. (2018). The neural correlates of Childhood Trauma Questionnaire scores in adults: A meta-analysis and review of functional magnetic resonance imaging studies. Development and psychopathology, 30(4), 1475–1485. https://doi.org/10.1017/S0954579417001717
Hein, T. C., & Monk, C. S. (2017). Research Review: Neural response to threat in children, adolescents, and adults after child maltreatment - a quantitative meta-analysis. Journal of child psychology and psychiatry, and allied disciplines, 58(3), 222–230. https://doi.org/10.1111/jcpp.12651
Ho, T. C., Gutman, B., Pozzi, E., Grabe, H. J., Hosten, N., Wittfeld, K., Völzke, H., Baune, B., Dannlowski, U., Förster, K., Grotegerd, D., Redlich, R., Jansen, A., Kircher, T., Krug, A., Meinert, S., Nenadic, I., Opel, N., Dinga, R., Veltman, D. J., … Schmaal, L. (2022). Subcortical shape alterations in major depressive disorder: Findings from the ENIGMA major depressive disorder working group. Human brain mapping, 43(1), 341–351. https://doi.org/10.1002/hbm.24988
Holz, N. E., Berhe, O., Sacu, S., Schwarz, E., Tesarz, J., Heim, C. M., & Tost, H. (2023). Early Social Adversity, Altered Brain Functional Connectivity, and Mental Health. Biological psychiatry, 93(5), 430–441. https://doi.org/10.1016/j.biopsych.2022.10.019
Holz, N. E., Boecker-Schlier, R., Buchmann, A. F., Blomeyer, D., Jennen-Steinmetz, C., Baumeister, S., Plichta, M. M., Cattrell, A., Schumann, G., Esser, G., Schmidt, M., Buitelaar, J., Meyer-Lindenberg, A., Banaschewski, T., Brandeis, D., & Laucht, M. (2017). Ventral striatum and amygdala activity as convergence sites for early adversity and conduct disorder. Social cognitive and affective neuroscience, 12(2), 261–272. https://doi.org/10.1093/scan/nsw120
Houtepen, L. C., Hardy, R., Maddock, J., Kuh, D., Anderson, E. L., Relton, C. L., Suderman, M. J., & Howe, L. D. (2018). Childhood adversity and DNA methylation in two population-based cohorts. Translational psychiatry, 8(1), 266. https://doi.org/10.1038/s41398-018-0307-3
Ibrahim, P., Almeida, D., Nagy, C., & Turecki, G. (2021). Molecular impacts of childhood abuse on the human brain. Neurobiology of stress, 15, 100343. https://doi.org/10.1016/j.ynstr.2021.100343
Kaiser, R. H., Andrews-Hanna, J. R., Wager, T. D., & Pizzagalli, D. A. (2015). Large-Scale Network Dysfunction in Major Depressive Disorder: A Meta-analysis of Resting-State Functional Connectivity. JAMA psychiatry, 72(6), 603–611. https://doi.org/10.1001/jamapsychiatry.2015.0071
Khoury, J. E., Bosquet Enlow, M., Plamondon, A., & Lyons-Ruth, K. (2019). The association between adversity and hair cortisol levels in humans: A meta-analysis. Psychoneuroendocrinology, 103, 104–117. https://doi.org/10.1016/j.psyneuen.2019.01.009
Klengel, T., Mehta, D., Anacker, C., Rex-Haffner, M., Pruessner, J. C., Pariante, C. M., Pace, T. W., Mercer, K. B., Mayberg, H. S., Bradley, B., Nemeroff, C. B., Holsboer, F., Heim, C. M., Ressler, K. J., Rein, T., & Binder, E. B. (2013). Allele-specific FKBP5 DNA demethylation mediates gene-childhood trauma interactions. Nature neuroscience, 16(1), 33–41. https://doi.org/10.1038/nn.3275
Koch, S. B., van Zuiden, M., Nawijn, L., Frijling, J. L., Veltman, D. J., & Olff, M. (2016). ABERRANT RESTING-STATE BRAIN ACTIVITY IN POSTTRAUMATIC STRESS DISORDER: A META-ANALYSIS AND SYSTEMATIC REVIEW. Depression and anxiety, 33(7), 592–605. https://doi.org/10.1002/da.22478
Korosi, A., & Baram, T. Z. (2009). The pathways from mother's love to baby's future. Frontiers in behavioral neuroscience, 3, 27. https://doi.org/10.3389/neuro.08.027.2009
Kraaijenvanger, E. J., Pollok, T. M., Monninger, M., Kaiser, A., Brandeis, D., Banaschewski, T., & Holz, N. E. (2020). Impact of early life adversities on human brain functioning: A coordinate-based meta-analysis. Neuroscience and biobehavioral reviews, 113, 62–76. https://doi.org/10.1016/j.neubiorev.2020.03.008
Kraynak, T. E., Marsland, A. L., Hanson, J. L., & Gianaros, P. J. (2019). Retrospectively reported childhood physical abuse, systemic inflammation, and resting corticolimbic connectivity in midlife adults. Brain, behavior, and immunity, 82, 203–213. https://doi.org/10.1016/j.bbi.2019.08.186
Kuhlman, K. R., Horn, S. R., Chiang, J. J., & Bower, J. E. (2020). Early life adversity exposure and circulating markers of inflammation in children and adolescents: A systematic review and meta-analysis. Brain, behavior, and immunity, 86, 30–42. https://doi.org/10.1016/j.bbi.2019.04.028
Labonté, B., Azoulay, N., Yerko, V., Turecki, G., & Brunet, A. (2014). Epigenetic modulation of glucocorticoid receptors in posttraumatic stress disorder. Translational psychiatry, 4(3), e368. https://doi.org/10.1038/tp.2014.3
Lamers, F., Vogelzangs, N., Merikangas, K. R., de Jonge, P., Beekman, A. T., & Penninx, B. W. (2013). Evidence for a differential role of HPA-axis function, inflammation and metabolic syndrome in melancholic versus atypical depression. Molecular psychiatry, 18(6), 692–699. https://doi.org/10.1038/mp.2012.144
Lecei, A., & van Winkel, R. (2020). Hippocampal pattern separation of emotional information determining risk or resilience in individuals exposed to childhood trauma: Linking exposure to neurodevelopmental alterations and threat anticipation. Neuroscience and biobehavioral reviews, 108, 160–170. https://doi.org/10.1016/j.neubiorev.2019.11.010
Levy-Gigi, E., Szabó, C., Kelemen, O., & Kéri, S. (2013). Association among clinical response, hippocampal volume, and FKBP5 gene expression in individuals with posttraumatic stress disorder receiving cognitive behavioral therapy. Biological psychiatry, 74(11), 793–800. https://doi.org/10.1016/j.biopsych.2013.05.017
Lim, L., Howells, H., Radua, J., & Rubia, K. (2020). Aberrant structural connectivity in childhood maltreatment: A meta-analysis. Neuroscience and biobehavioral reviews, 116, 406–414. https://doi.org/10.1016/j.neubiorev.2020.07.004
Liston, C., McEwen, B. S., & Casey, B. J. (2009). Psychosocial stress reversibly disrupts prefrontal processing and attentional control. Proceedings of the National Academy of Sciences of the United States of America, 106(3), 912–917. https://doi.org/10.1073/pnas.0807041106
Luijten, M., Schellekens, A. F., Kühn, S., Machielse, M. W., & Sescousse, G. (2017). Disruption of Reward Processing in Addiction : An Image-Based Meta-analysis of Functional Magnetic Resonance Imaging Studies. JAMA psychiatry, 74(4), 387–398. https://doi.org/10.1001/jamapsychiatry.2016.3084
Lutz, P. E., Gross, J. A., Dhir, S. K., Maussion, G., Yang, J., Bramoulle, A., Meaney, M. J., & Turecki, G. (2018). Epigenetic Regulation of the Kappa Opioid Receptor by Child Abuse. Biological psychiatry, 84(10), 751–761. https://doi.org/10.1016/j.biopsych.2017.07.012
Lutz, P. E., & Turecki, G. (2014). DNA methylation and childhood maltreatment: from animal models to human studies. Neuroscience, 264, 142–156. https://doi.org/10.1016/j.neuroscience.2013.07.069
Maes, M., Rachayon, M., Jirakran, K., Sodsai, P., Klinchanhom, S., Debnath, M., Basta-Kaim, A., Kubera, M., Almulla, A. F., & Sughondhabirom, A. (2022). Adverse Childhood Experiences Predict the Phenome of Affective Disorders and These Effects Are Mediated by Staging, Neuroimmunotoxic and Growth Factor Profiles. Cells, 11(9), 1564. https://doi.org/10.3390/cells11091564
Makinodan, M., Rosen, K. M., Ito, S., & Corfas, G. (2012). A critical period for social experience-dependent oligodendrocyte maturation and myelination. Science (New York, N.Y.), 337(6100), 1357–1360. https://doi.org/10.1126/science.1220845
Maras, P. M., & Baram, T. Z. (2012). Sculpting the hippocampus from within: stress, spines, and CRH. Trends in neurosciences, 35(5), 315–324. https://doi.org/10.1016/j.tins.2012.01.005
Marzi, S. J., Sugden, K., Arseneault, L., Belsky, D. W., Burrage, J., Corcoran, D. L., Danese, A., Fisher, H. L., Hannon, E., Moffitt, T. E., Odgers, C. L., Pariante, C., Poulton, R., Williams, B. S., Wong, C. C. Y., Mill, J., & Caspi, A. (2018). Analysis of DNA Methylation in Young People: Limited Evidence for an Association Between Victimization Stress and Epigenetic Variation in Blood. The American journal of psychiatry, 175(6), 517–529. https://doi.org/10.1176/appi.ajp.2017.17060693
Matosin, N., Halldorsdottir, T., & Binder, E. B. (2018). Understanding the Molecular Mechanisms Underpinning Gene by Environment Interactions in Psychiatric Disorders: The FKBP5 Model. Biological psychiatry, 83(10), 821–830. https://doi.org/10.1016/j.biopsych.2018.01.021
Mattson, W. I., Hyde, L. W., Shaw, D. S., Forbes, E. E., & Monk, C. S. (2016). Clinical neuroprediction: Amygdala reactivity predicts depressive symptoms 2 years later. Social cognitive and affective neuroscience, 11(6), 892–898. https://doi.org/10.1093/scan/nsw018
McGowan, P. O., Sasaki, A., D'Alessio, A. C., Dymov, S., Labonté, B., Szyf, M., Turecki, G., & Meaney, M. J. (2009). Epigenetic regulation of the glucocorticoid receptor in human brain associates with childhood abuse. Nature neuroscience, 12(3), 342–348. https://doi.org/10.1038/nn.2270
Mehta, M. A., Gore-Langton, E., Golembo, N., Colvert, E., Williams, S. C., & Sonuga-Barke, E. (2010). Hyporesponsive reward anticipation in the basal ganglia following severe institutional deprivation early in life. Journal of cognitive neuroscience, 22(10), 2316–2325. https://doi.org/10.1162/jocn.2009.21394
Mian, O., Belsky, D. W., Cohen, A. A., Anderson, L. N., Gonzalez, A., Ma, J., Sloboda, D. M., Bowdish, D. M., & Verschoor, C. P. (2022). Associations between exposure to adverse childhood experiences and biological aging: Evidence from the Canadian Longitudinal Study on Aging. Psychoneuroendocrinology, 142, 105821. https://doi.org/10.1016/j.psyneuen.2022.105821
Miller, G. E., Chen, E., & Zhou, E. S. (2007). If it goes up, must it come down? Chronic stress and the hypothalamic-pituitary-adrenocortical axis in humans. Psychological bulletin, 133(1), 25–45. https://doi.org/10.1037/0033-2909.133.1.25
Morris, M. C., Compas, B. E., & Garber, J. (2012). Relations among posttraumatic stress disorder, comorbid major depression, and HPA function: a systematic review and meta-analysis. Clinical psychology review, 32(4), 301–315. https://doi.org/10.1016/j.cpr.2012.02.002
Mothersill, O., & Donohoe, G. (2016). Neural effects of social environmental stress - an activation likelihood estimation meta-analysis. Psychological medicine, 46(10), 2015–2023. https://doi.org/10.1017/S0033291716000477
Murgatroyd, C., Quinn, J. P., Sharp, H. M., Pickles, A., & Hill, J. (2015). Effects of prenatal and postnatal depression, and maternal stroking, at the glucocorticoid receptor gene. Translational psychiatry, 5(5), e560. https://doi.org/10.1038/tp.2014.140
Nakamoto, K., & Tokuyama, S. (2023). Stress-Induced Changes in the Endogenous Opioid System Cause Dysfunction of Pain and Emotion Regulation. International journal of molecular sciences, 24(14), 11713. https://doi.org/10.3390/ijms241411713
Nelson, M. D., & Tumpap, A. M. (2017). Posttraumatic stress disorder symptom severity is associated with left hippocampal volume reduction: a meta-analytic study. CNS spectrums, 22(4), 363–372. https://doi.org/10.1017/S1092852916000833
Olson, A. K., Eadie, B. D., Ernst, C., & Christie, B. R. (2006). Environmental enrichment and voluntary exercise massively increase neurogenesis in the adult hippocampus via dissociable pathways. Hippocampus, 16(3), 250–260. https://doi.org/10.1002/hipo.20157
Orso, R., Creutzberg, K. C., Kestering-Ferreira, E., Wearick-Silva, L. E., Tractenberg, S. G., & Grassi-Oliveira, R. (2020). Maternal Separation Combined With Limited Bedding Increases Anxiety-Like Behavior and Alters Hypothalamic-Pituitary-Adrenal Axis Function of Male BALB/cJ Mice. Frontiers in behavioral neuroscience, 14, 600766. https://doi.org/10.3389/fnbeh.2020.600766
Parent, J., Parade, S. H., Laumann, L. E., Ridout, K. K., Yang, B. Z., Marsit, C. J., Seifer, R., & Tyrka, A. R. (2017). Dynamic stress-related epigenetic regulation of the glucocorticoid receptor gene promoter during early development: The role of child maltreatment. Development and psychopathology, 29(5), 1635–1648. https://doi.org/10.1017/S0954579417001298
Philip, N. S., Sweet, L. H., Tyrka, A. R., Price, L. H., Bloom, R. F., & Carpenter, L. L. (2013). Decreased default network connectivity is associated with early life stress in medication-free healthy adults. European neuropsychopharmacology : the journal of the European College of Neuropsychopharmacology, 23(1), 24–32. https://doi.org/10.1016/j.euroneuro.2012.10.008
Pizzagalli D. A. (2022). Toward a Better Understanding of the Mechanisms and Pathophysiology of Anhedonia: Are We Ready for Translation?. The American journal of psychiatry, 179(7), 458–469. https://doi.org/10.1176/appi.ajp.20220423
Plotsky, P. M., & Meaney, M. J. (1993). Early, postnatal experience alters hypothalamic corticotropin-releasing factor (CRF) mRNA, median eminence CRF content and stress-induced release in adult rats. Brain research. Molecular brain research, 18(3), 195–200. https://doi.org/10.1016/0169-328x(93)90189-v
Priest, N., Guo, S., Gondek, D., Lacey, R. E., Burgner, D., Downes, M., Slopen, N., Goldfeld, S., Moreno-Betancur, M., Kerr, J. A., Cahill, S., Wake, M., Juonala, M., Lycett, K., & O'Connor, M. (2022). The effect of adverse and positive experiences on inflammatory markers in Australian and UK children. Brain, behavior, & immunity - health, 26, 100550. https://doi.org/10.1016/j.bbih.2022.100550
Reid, B. M., Doom, J. R., Argote, R. B., Correa-Burrows, P., Lozoff, B., Blanco, E., & Gahagan, S. (2020). Pathways to inflammation in adolescence through early adversity, childhood depressive symptoms, and body mass index: A prospective longitudinal study of Chilean infants. Brain, behavior, and immunity, 86, 4–13. https://doi.org/10.1016/j.bbi.2019.06.003
Riem, M. M., Alink, L. R., Out, D., Van Ijzendoorn, M. H., & Bakermans-Kranenburg, M. J. (2015). Beating the brain about abuse: Empirical and meta-analytic studies of the association between maltreatment and hippocampal volume across childhood and adolescence. Development and psychopathology, 27(2), 507–520. https://doi.org/10.1017/S0954579415000127
Ronaldson, A., Gazali, A. M., Zalli, A., Kaiser, F., Thompson, S. J., Henderson, B., Steptoe, A., & Carvalho, L. (2016). Increased percentages of regulatory T cells are associated with inflammatory and neuroendocrine responses to acute psychological stress and poorer health status in older men and women. Psychopharmacology, 233(9), 1661–1668. https://doi.org/10.1007/s00213-015-3876-3
Roozendaal, B., McEwen, B. S., & Chattarji, S. (2009). Stress, memory and the amygdala. Nature reviews. Neuroscience, 10(6), 423–433. https://doi.org/10.1038/nrn2651
Ross, M. C., Heilicher, M., & Cisler, J. M. (2021). Functional imaging correlates of childhood trauma: A qualitative review of past research and emerging trends. Pharmacology, biochemistry, and behavior, 211, 173297. https://doi.org/10.1016/j.pbb.2021.173297
Rutter M. (1998). Developmental catch-up, and deficit, following adoption after severe global early privation. English and Romanian Adoptees (ERA) Study Team. Journal of child psychology and psychiatry, and allied disciplines, 39(4), 465–476.
Sánchez, M. M., Noble, P. M., Lyon, C. K., Plotsky, P. M., Davis, M., Nemeroff, C. B., & Winslow, J. T. (2005). Alterations in diurnal cortisol rhythm and acoustic startle response in nonhuman primates with adverse rearing. Biological psychiatry, 57(4), 373–381. https://doi.org/10.1016/j.biopsych.2004.11.032
Sanders, J., & Nemeroff, C. (2016). The CRF System as a Therapeutic Target for Neuropsychiatric Disorders. Trends in pharmacological sciences, 37(12), 1045–1054. https://doi.org/10.1016/j.tips.2016.09.004
Sapolsky R. M. (1996). Stress, Glucocorticoids, and Damage to the Nervous System: The Current State of Confusion. Stress (Amsterdam, Netherlands), 1(1), 1–19. https://doi.org/10.3109/10253899609001092
Sarapas, C., Cai, G., Bierer, L. M., Golier, J. A., Galea, S., Ising, M., Rein, T., Schmeidler, J., Müller-Myhsok, B., Uhr, M., Holsboer, F., Buxbaum, J. D., & Yehuda, R. (2011). Genetic markers for PTSD risk and resilience among survivors of the World Trade Center attacks. Disease markers, 30(2-3), 101–110. https://doi.org/10.3233/DMA-2011-0764
Schär, S., Mürner-Lavanchy, I., Schmidt, S. J., Koenig, J., & Kaess, M. (2022). Child maltreatment and hypothalamic-pituitary-adrenal axis functioning: A systematic review and meta-analysis. Frontiers in neuroendocrinology, 66, 100987. https://doi.org/10.1016/j.yfrne.2022.100987
Schechter, D. S., Moser, D. A., Paoloni-Giacobino, A., Stenz, L., Gex-Fabry, M., Aue, T., Adouan, W., Cordero, M. I., Suardi, F., Manini, A., Sancho Rossignol, A., Merminod, G., Ansermet, F., Dayer, A. G., & Rusconi Serpa, S. (2015). Methylation of NR3C1 is related to maternal PTSD, parenting stress and maternal medial prefrontal cortical activity in response to child separation among mothers with histories of violence exposure. Frontiers in psychology, 6, 690. https://doi.org/10.3389/fpsyg.2015.00690
Schmaal, L., Veltman, D. J., van Erp, T. G., Sämann, P. G., Frodl, T., Jahanshad, N., Loehrer, E., Tiemeier, H., Hofman, A., Niessen, W. J., Vernooij, M. W., Ikram, M. A., Wittfeld, K., Grabe, H. J., Block, A., Hegenscheid, K., Völzke, H., Hoehn, D., Czisch, M., Lagopoulos, J., … Hibar, D. P. (2016). Subcortical brain alterations in major depressive disorder: findings from the ENIGMA Major Depressive Disorder working group. Molecular psychiatry, 21(6), 806–812. https://doi.org/10.1038/mp.2015.69
Schulze, L., Schulze, A., Renneberg, B., Schmahl, C., & Niedtfeld, I. (2019). Neural Correlates of Affective Disturbances: A Comparative Meta-analysis of Negative Affect Processing in Borderline Personality Disorder, Major Depressive Disorder, and Posttraumatic Stress Disorder. Biological psychiatry. Cognitive neuroscience and neuroimaging, 4(3), 220–232. https://doi.org/10.1016/j.bpsc.2018.11.004
Shafie, M., Shahmohamadi, E., Cattarinussi, G., Sanjari Moghaddam, H., Akhondzadeh, S., Sambataro, F., Moltrasio, C., & Delvecchio, G. (2023). Resting-state functional magnetic resonance imaging alterations in borderline personality disorder: A systematic review. Journal of affective disorders, 341, 335–345. https://doi.org/10.1016/j.jad.2023.09.001
Sheerin, C. M., Lind, M. J., Bountress, K. E., Marraccini, M. E., Amstadter, A. B., Bacanu, S. A., & Nugent, N. R. (2020). Meta-Analysis of Associations Between Hypothalamic-Pituitary-Adrenal Axis Genes and Risk of Posttraumatic Stress Disorder. Journal of traumatic stress, 33(5), 688–698. https://doi.org/10.1002/jts.22484
Shirtcliff, E. A., Hanson, J. L., Ruttle, P. L., Smith, B., & Pollak, S. D. (2024). Cortisol's diurnal rhythm indexes the neurobiological impact of child adversity in adolescence. Biological psychology, 187, 108766. https://doi.org/10.1016/j.biopsycho.2024.108766
Slopen, N., Kubzansky, L. D., McLaughlin, K. A., & Koenen, K. C. (2013). Childhood adversity and inflammatory processes in youth: a prospective study. Psychoneuroendocrinology, 38(2), 188–200. https://doi.org/10.1016/j.psyneuen.2012.05.013
Takahashi, Y., Kubo, R., Sano, R., Kuninaka, H., Murayama, M., Hayakawa, A., & Kominato, Y. (2018). DNA methylation of the NR3C1 promoter region in brains of pediatric victims of physical abuse. Neurocase, 24(5-6), 269–275. https://doi.org/10.1080/13554794.2019.1582678
Tanti, A., Kim, J. J., Wakid, M., Davoli, M. A., Turecki, G., & Mechawar, N. (2018). Child abuse associates with an imbalance of oligodendrocyte-lineage cells in ventromedial prefrontal white matter. Molecular psychiatry, 23(10), 2018–2028. https://doi.org/10.1038/mp.2017.231
Teicher, M. H., & Samson, J. A. (2013). Childhood maltreatment and psychopathology: A case for ecophenotypic variants as clinically and neurobiologically distinct subtypes. The American journal of psychiatry, 170(10), 1114–1133. https://doi.org/10.1176/appi.ajp.2013.12070957
Thomas, N., Gurvich, C., Hudaib, A. R., Gavrilidis, E., & Kulkarni, J. (2019). Systematic review and meta-analysis of basal cortisol levels in Borderline Personality Disorder compared to non-psychiatric controls. Psychoneuroendocrinology, 102, 149–157. https://doi.org/10.1016/j.psyneuen.2018.12.009
Trickett, P. K., Noll, J. G., Susman, E. J., Shenk, C. E., & Putnam, F. W. (2010). Attenuation of cortisol across development for victims of sexual abuse. Development and psychopathology, 22(1), 165–175. https://doi.org/10.1017/S0954579409990332
Ulrich-Lai, Y. M., & Herman, J. P. (2009). Neural regulation of endocrine and autonomic stress responses. Nature reviews. Neuroscience, 10(6), 397–409. https://doi.org/10.1038/nrn2647
Van Hulle, C. A., Shirtcliff, E. A., Lemery-Chalfant, K., & Goldsmith, H. H. (2012). Genetic and environmental influences on individual differences in cortisol level and circadian rhythm in middle childhood. Hormones and behavior, 62(1), 36–42. https://doi.org/10.1016/j.yhbeh.2012.04.014
van Oers, H. J., de Kloet, E. R., Whelan, T., & Levine, S. (1998). Maternal deprivation effect on the infant's neural stress markers is reversed by tactile stimulation and feeding but not by suppressing corticosterone. The Journal of neuroscience : the official journal of the Society for Neuroscience, 18(23), 10171–10179. https://doi.org/10.1523/JNEUROSCI.18-23-10171.1998
van Zuiden, M., Kavelaars, A., Geuze, E., Olff, M., & Heijnen, C. J. (2013). Predicting PTSD: pre-existing vulnerabilities in glucocorticoid-signaling and implications for preventive interventions. Brain, behavior, and immunity, 30, 12–21. https://doi.org/10.1016/j.bbi.2012.08.015
Veenema A. H. (2009). Early life stress, the development of aggression and neuroendocrine and neurobiological correlates: what can we learn from animal models?. Frontiers in neuroendocrinology, 30(4), 497–518. https://doi.org/10.1016/j.yfrne.2009.03.003
Vyas, N., Wimberly, C. E., Beaman, M. M., Kaplan, S. J., Rasmussen, L. J. H., Wertz, J., Gifford, E. J., & Walsh, K. M. (2023). Systematic review and meta-analysis of the effect of adverse childhood experiences (ACEs) on brain-derived neurotrophic factor (BDNF) levels. Psychoneuroendocrinology, 151, 106071. https://doi.org/10.1016/j.psyneuen.2023.106071
Wadji, D. L., Tandon, T., Ketcha Wanda, G. J. M., Wicky, C., Dentz, A., Hasler, G., Morina, N., & Martin-Soelch, C. (2021). Child maltreatment and NR3C1 exon 1F methylation, link with deregulated hypothalamus-pituitary-adrenal axis and psychopathology: A systematic review. Child abuse & neglect, 122, 105304. https://doi.org/10.1016/j.chiabu.2021.105304
Wang, Q., Shelton, R. C., & Dwivedi, Y. (2018). Interaction between early-life stress and FKBP5 gene variants in major depressive disorder and post-traumatic stress disorder: A systematic review and meta-analysis. Journal of affective disorders, 225, 422–428. https://doi.org/10.1016/j.jad.2017.08.066
Watt, T., Ceballos, N., Kim, S., Pan, X., & Sharma, S. (2019). The Unique Nature of Depression and Anxiety among College Students with Adverse Childhood Experiences. Journal of child & adolescent trauma, 13(2), 163–172. https://doi.org/10.1007/s40653-019-00270-4
Weaver, I. C., Cervoni, N., Champagne, F. A., D'Alessio, A. C., Sharma, S., Seckl, J. R., Dymov, S., Szyf, M., & Meaney, M. J. (2004). Epigenetic programming by maternal behavior. Nature neuroscience, 7(8), 847–854. https://doi.org/10.1038/nn1276
Wilker, S., Pfeiffer, A., Kolassa, S., Elbert, T., Lingenfelder, B., Ovuga, E., Papassotiropoulos, A., de Quervain, D., & Kolassa, I. T. (2014). The role of FKBP5 genotype in moderating long-term effectiveness of exposure-based psychotherapy for posttraumatic stress disorder. Translational psychiatry, 4(6), e403. https://doi.org/10.1038/tp.2014.49
Yehuda, R., Flory, J. D., Bierer, L. M., Henn-Haase, C., Lehrner, A., Desarnaud, F., Makotkine, I., Daskalakis, N. P., Marmar, C. R., & Meaney, M. J. (2015). Lower methylation of glucocorticoid receptor gene promoter 1F in peripheral blood of veterans with posttraumatic stress disorder. Biological psychiatry, 77(4), 356–364. https://doi.org/10.1016/j.biopsych.2014.02.006
Young, E. S., Doom, J. R., Farrell, A. K., Carlson, E. A., Englund, M. M., Miller, G. E., Gunnar, M. R., Roisman, G. I., & Simpson, J. A. (2021). Life stress and cortisol reactivity: An exploratory analysis of the effects of stress exposure across life on HPA-axis functioning. Development and psychopathology, 33(1), 301–312. https://doi.org/10.1017/S0954579419001779